Location: Home >> Detail
Immunometabolism. 2020;2(2):e200017. https://doi.org/10.20900/immunometab20200017

1 School of Medicine, Stanford University, Stanford, CA 94305, USA
2 Department of Medicine, Palo Alto Veterans Administration Healthcare System, Palo Alto, CA 94304, USA
* Correspondence: Cornelia M. Weyand, Tel.: +1-650-723-9027; Fax: +1-650-721-1251.
This article belongs to the Virtual Special Issue "Immunometabolism in Autoimmune Diseases"
Rheumatoid arthritis (RA) and systemic lupus erythematosus (SLE) are relatively common autoimmune diseases, often considered prototypic examples for how protective immunity switches to destructive immunity. The autoantigens recognized in RA and SLE are distinct, clinical manifestations are partially overlapping. A shared feature is the propensity of the adaptive immune system to respond inappropriately, with T cell hyper-responsiveness a pinnacle pathogenic defect. Upon antigen recognition, T cells mobilize a multi-pranged metabolic program, enabling them to massively expand and turn into highly mobile effector cells. Current evidence supports that T cells from patients with RA or SLE adopt metabolic programs different from healthy T cells, in line with the concept that autoimmune effector functions rely on specified pathways of energy sensing, energy generation and energy utilization. Due to misrouting of the energy sensor AMPK, RA T cells have a defect in balancing catabolic and anabolic processes and deviate towards a cell-building program. They supply biosynthetic precursors by shunting glucose away from glycolytic breakdown towards the pentose phosphate pathway and upregulate lipogenesis, enabling cellular motility and tissue invasiveness. Conversely, T cells from SLE patients are committed to high glycolytic flux, overusing the mitochondrial machinery and imposing oxidative stress. Typically, disease-relevant effector functions in SLE are associated with inappropriate activation of the key metabolic regulator mTORC1. Taken together, disease-specific metabolic signatures in RA and SLE represent vulnerabilities that are therapeutically targetable to suppress pathogenic immune responses.
Rheumatoid arthritis (RA) and systemic lupus erythematosus (SLE) are two representative autoimmune diseases that affect large patient populations and display typical features of immune system abnormalities. Both diseases can cause a broad spectrum of organ pathology. In RA, the main manifestation of auto-aggression is a chronic-persistent, symmetrical and destructive joint inflammation, while patients affected by SLE can have inflammatory injury of almost every organ system. RA and SLE share in common that affected individuals have a multi-fold higher risk to develop cardiovascular complications [1–3]; often understood as a complication of chronic-smoldering inflammation. Cardiovascular risk imposed by long-standing autoimmunity suggests that deficiencies of tissue homeostasis and repair may be part of the overall immune defect.
For most autoimmune diseases, the breakdown of self-tolerance and autoantibody production precedes clinical onset of disease by years to decades [4,5], indicating a fundamental and chronic rewiring of the immune system. HLA class II-restricted CD4 T cells, major players in inflammatory lesions and indispensable to induce and sustain autoantibody production by B cells, are central drivers in autoimmune diseases [6]. T cells rely strongly on bio-energetic and biosynthetic plasticity to fulfill their functions. With the ability to massively proliferate and commit to lineage differentiation, to invade and reside in different tissue microenvironments, such T cells encounter heterogeneous metabolic demands. Thus, the metabolic machinery is vulnerable to even minor changes translating into major, unexpected outcomes. In this review, we will summarize metabolic features of T cells in both RA and SLE patients; identifying metabolic checkpoints where T cells are mistakenly led into distinct disease-promoting directions.
With fatty acids as the main fuel during the naïve state [7], effector T cells exhibit a dramatic metabolic switch towards glycolysis upon activation, upregulating glucose transporters and a large spectrum of glycolytic enzymes [8,9]. In contrast, regulatory T cells and memory T cells depend mostly on fatty acid oxidation [7,10] to support their survival needs and their functional activities. As a rule, maintenance of T cell nativity, memory and regulatory function is closely linked to lipid utilization and thus is ultimately dependent on mitochondrial fitness [11], whereas effector T cells, including the pro-inflammatory T cells that sustain tissue inflammation, basically, fuel their energy demands through fast access to glucose and glycolysis.
Glucose is actively transported into the cell and is catalyzed into glucose-6-phosphate (G6P) by hexokinases (HKs). Subsequently, G6P either goes into glycolytic breakdown to produce pyruvate or enters the pentose phosphate pathway to generate NADPH and nucleotide precursors. A key event in the glycolytic breakdown is the phosphorylation of fructose 6phosphate to fructose-1, 6-bisphosphate through 6phosphofructo1kinase (PFK1), an irreversible reaction which commits glucose to glycolysis. PFK1 activity is dominantly controlled by its allosteric activator fructose-2, 6bisphosphate (F2, 6BP), generated by 6phosphofructo2kinase/ fructose2, 6bis-phosphatase-3 (PFKFB3). Hence, PFKFB3 critically regulates the glycolytic rate and PFKFB3 is the target of regulatory events that dictate the efficiency of cytoplasmic glucose utilization. T cells from RA patients fail to upregulate PFKFB3 upon activation (Figure 1), suppressing glycolytic breakdown and limiting pyruvate/lactate production [12,13]. On the contrary, RA T cells transcriptionally upregulate G6PD, shifting glucose into the pentose phosphate pathway (PPP) (Figure 1). The outcome of a dysbalance in the PFK/G6PD ratio is the overproduction of NADPH and accumulation of reduced glutathione, creating a reductive intracellular environment [14,15]. This fundamental change in redox homeostasis significantly disrupts proper sensing of the energy state, mistakenly promoting cell proliferation because of insufficient activation of the cell cycle checkpoint kinase ATM [12,16]. Most of these data have been collected in patient-derived CD4 T cells and the metabolic switch from catabolism and glucose breakdown to anabolism and generation of biosynthetic precursor is already present in the naïve CD4 T cell population, and these metabolic features are specific for the T cell compartment [17]. It is therefore highly unlikely that these metabolic adaptations are the result of T cells living in an inflammatory tissue environment. Rather, the invasion of T cells into peripheral tissue sites, where they drive inflammatory pathways and set up tissue residence, is already a consequence of metabolic reprogramming.
Once CD4 T cells in RA patients have fully committed to effector cell differentiation, they utilize metabolic programs typical for effector T cells (reviewed in [7–11]).
The inflamed synovial lesions exclusively contain memory and effector T cells and these cells are exposed to metabolic cues imposed by the tissue environment. Besides having to share energy resources, lesional T cells will receive metabolic signals from neighboring cell populations, which are highly metabolically active [18]. A dominant metabolite encountered in the inflamed joint is lactate [19,20], a breakdown product of glucose, produced by metabolically active stromal cells, endothelial cells and invading immune cells. Lactate has been implicated in directly shaping T cell effector functions by increased IL-17 production via nuclear PKM2/STAT3 and enhanced fatty acid synthesis [21]. Also, lactate can inhibit the migration of both CD4 and CD8 T cells, thus causing T cell entrapment within the inflamed synovial tissue [22]. Elevated frequencies of ICOS+ Tfh cells have been described in the synovial fluid of RA patients, and targeting glycolysis in such Tfh cells has been reported to ameliorate established collagen-induced arthritis [23].
Fundamentally different from RA T cells, CD4 T cells from lupus patients and lupus-prone mice display high demand for glucose [24,25] and depend on rapid production of ATP, requiring both mitochondrial activity and cytoplasmic glycolysis [26]. Interestingly, in SLE T cells the pentose phosphate pathway appears to be also hyperactive, although the underlying mechanisms remain unclear [27].
Much of the focus in SLE has been on follicular helper CD4 T (Tfh) cells, which are critically involved in providing help for B cell maturation and autoantibodies production. In SLE patients, frequencies of circulating Tfh cells are elevated and such frequencies are often correlated with disease load [28]. Metabolic needs of autoreactive Tfh cells appear to be different from influenza virus-specific helper T cells [29]. In murine models of SLE, broad inhibition of glycolysis, achieved by treating with 2-deoxyglucose (2-DG), successfully suppressed the expansion of autoreactive Tfh cells, but seemed to not affect B cell responses to exogenous antigens nor the induction of anti-influenza Tfh cells. These findings suggest differential metabolic wiring and differential sensitivity to intervention of protective and tissue-inflammatory T cells.
Amongst T cell effector cells, Th17 cells stand out as pathogenic drivers in SLE. Patients with SLE have a higher frequency of Th17 cells, which contribute to the establishment of proinflammatory conditions by infiltrating multiple organs [30,31]. Therefore, the molecular machinery that induces Th17 differentiation is of high interest in the investigation of SLE pathogenesis. Inducible cAMP early repressor (ICER), an isoform of the cAMP Response Element Modulator (CREM), is exclusively induced in Th17 cells and promotes Th17 cell differentiation via binding to the IL-17 promoter region to recruit the canonical enhancer RORγt. Increased ICER concentrations in CD4 T cells from patients with SLE have been associated with Th17 cell differentiation and disease severity [32]. In recent studies, ICER has been implicated in yet another mechanism promoting Th17 differentiation, and thus enhancing risk for SLE [33]. Th17 cells favor glycolytic metabolism, preferentially using pyruvate for lactate generation instead of mitochondria-dependent oxidative phosphorylation. Pyruvate dehydrogenase (PDH) is the bifurcation enzyme controlling the balance between lactate production and oxidative phosphorylation, with its activity critically controlled by its phosphorylation status. Pyruvate dehydrogenase phosphatase (PDP) can dephosphorylate and activate PDH. In SLE T cells, high levels of ICER inhibit pyruvate dehydrogenase phosphatase catalytic subunit 2 (PDP2) expression, impairing PDH dephosphorylation and activation, ultimately funneling glycolytic breakdown of glucose towards lactate (Figure 1). Forced overexpression of PDP2 reduced Th17 differentiation in CD4 T cells from both SLE patients and lupus-prone mice. These elegant studies have given new insights into regulatory functions of ICER, and how it targets the metabolic machinery. ICER’s downstream targets PDP2/PDH are obvious checkpoints in Th17 differentiation and are possibly druggable to avoid the Th17 bias driving SLE pathology (Figure 1).
 Figure 1. Rerouting of glucose utilization in RA and SLE T cells. RA T cells fail to upregulate PFKFB3, a key enzyme in the glycolytic pathway, leading to low production of pyruvate and lactate. Instead, RA T cells upregulate expression and function of G6PD, the enzyme that controls the entry to the pentose phosphate pathway (PPP). As a result, RA T cells produce more NADPH and shift the cellular redox balance towards a reductive environment (low ROS, high glutathione). T cells from SLE patients increase glycolytic breakdown of glucose and produce copious amounts of pyruvate and lactate. The intracellular environment is biased towards oxidative stress. One mechanism underlying the preferential routing of glucose towards glycolysis relates to the suppression of pyruvate dehydrogenase (PDH). An upstream event is the high expression of the transcription factor inducible cAMP early repressor (ICER), which, in turn, leads to inhibition of PDH, and suppression of PDH. ICER is considered a lineage-promoting transcription factor promoting Th17 commitment. Th17 cells are recognized as key effector cells in SLE. Overall, RA T cells are programmed towards anabolic activity, whereas SLE T cells favor catabolic metabolism. PPP, pentose phosphate pathway; ROS, reactive oxygen species; PDP2, pyruvate dehydrogenase phosphatase catalytic subunit 2.
Figure 1. Rerouting of glucose utilization in RA and SLE T cells. RA T cells fail to upregulate PFKFB3, a key enzyme in the glycolytic pathway, leading to low production of pyruvate and lactate. Instead, RA T cells upregulate expression and function of G6PD, the enzyme that controls the entry to the pentose phosphate pathway (PPP). As a result, RA T cells produce more NADPH and shift the cellular redox balance towards a reductive environment (low ROS, high glutathione). T cells from SLE patients increase glycolytic breakdown of glucose and produce copious amounts of pyruvate and lactate. The intracellular environment is biased towards oxidative stress. One mechanism underlying the preferential routing of glucose towards glycolysis relates to the suppression of pyruvate dehydrogenase (PDH). An upstream event is the high expression of the transcription factor inducible cAMP early repressor (ICER), which, in turn, leads to inhibition of PDH, and suppression of PDH. ICER is considered a lineage-promoting transcription factor promoting Th17 commitment. Th17 cells are recognized as key effector cells in SLE. Overall, RA T cells are programmed towards anabolic activity, whereas SLE T cells favor catabolic metabolism. PPP, pentose phosphate pathway; ROS, reactive oxygen species; PDP2, pyruvate dehydrogenase phosphatase catalytic subunit 2.
Telomeres serve as sensors of cellular aging and CD4 T cells from RA patients have age-inappropriate shortening of telomeres [34,35], indicating that RA T cells are prematurely aged [36]. Deficiencies of the nuclear DNA repair machinery have been demonstrated to play a critical role in this pre-mature aging phenotype [37,38]. Besides nuclear DNA damage, a recent study reported that defects in DNA damage repair of RA T cells extend to mitochondrial DNA (mtDNA) [39]. Molecular studies have identified a loss-of-function of MRE11A, a nuclease and key component of the MRN complex [40]. MRE11A downregulation results from transcriptional repression, but upstream signals inducing the MRE11Alow phenotype have not been identified yet. Functional studies have revealed that MRE11A localizes to the telomeric ends but is also placed in the mitochondria. In RA T cells, the nuclease is expressed at significantly lower concentrations, rendering both telomeric ends and mitochondrial DNA susceptible to unrepaired DNA damage [39,40]. Functional consequences include persistent damage repair activity and in the case of mitochondria, loss of function and leakage of damaged mtDNA into the cytoplasm (Figure 2). Mitochondrial biogenesis appears unaffected by the reduction of mitochondrial MRE11A, as healthy control and RA T cells carry similar mitochondrial mass. Specifically, mitochondrial oxygen consumption is suppressed in RA T cells, suggestive for a defect in the mitochondrial electron transport chain.
Also, insufficient repair and maintenance of mtDNA resulted in the leakage of unrepaired and oxidatively modified DNA into the cytoplasm, where it was recognized by DNA sensors and triggered AIM2 and NLRP3-mediated inflammasome assembly and activation [39] (Figure 2).
The concept of implicating inflammasome activation in T cells in chronic tissue inflammation is novel. T cell loss in HIV infection has been attributed to inflammasome-dependent pyroptotic cell death [41]. Whether the inflammasome shapes function and survival of SLE T cells is currently unexplored. However, inappropriate inflammasome activity in myeloid cells is considered a critical disease pathway in SLE [42]. Inflammasome-related IL-18/IL-1β signaling has been associated with abnormal T cell responses, especially the Th17/Treg balance [43], which represent a new paradigm in inflammatory diseases. Both IL-18 and IL-1β have been reported to enhance Th17 differentiation while inhibiting the regulatory function of Treg cells [44,45]. The NLRP3 inflammasome is hyperactive in macrophages of SLE patients [46], creating a pro-inflammatory tissue microenvironment. Similarly, inflammasome activation contributes to dendritic cell function and affects their role as antigen-presenting cells; e.g., by fostering T cell activation and Th1 and Th17 polarization [47]. The immediate products of inflammasome activation, IL-18 and IL-1β, have been positively associated with disease activity in SLE [48,49]. Opposing findings have been reported in systemic lupus nephritis, where inflammasome activation appears to suppress excessive immunity. Loss-of-function of the NLRP3/ASC inflammasome in C57BL/6-lpr/lpr mice enhanced dendritic cell and macrophage activation, triggered massive lymphoproliferation, increased T cell infiltrates in the lung and resulted in severe proliferative lupus nephritis [50]. These data suggest a homeostatic role of inflammasome activation, participating in balancing protective and destructive immunity. Underlying mechanisms are insufficiently understood, but suppression of TGF-β target genes may be involved [50].
A hallmark of CD4 T cells from SLE patients is mitochondrial hyperpolarization (MHP), measured as elevated mitochondrial transmembrane potential [ΔΨm] and reactive oxygen species (ROS) production [51,52] (Figure 2). In essence, lupus T cells are in a state of intense and lasting mitochondrial stimulation. Part of this increase in mitochondrial activity may be attributable to an increase in mitochondrial biogenesis [53], such that SLE T cells carry a higher load of mitochondria. Mitochondrial hyperactivity has numerous consequences for T cells, most prominently, the undermining of cellular defense mechanisms regulating oxidative stress. Glutathione, the most abundant intracellular antioxidant, is significantly reduced in SLE T cells, creating an oxidative intracellular environment. Multiple studies have corroborated that the mitochondrial building program is stimulated in SLE T cells. Large mitochondrial size, increased mitochondrial biogenesis and defective mitophagy are features of SLE T cells [53,54]. Upstream signals triggering the abnormal mitochondrial activation program remain elusive.
Downstream consequences of chronic mitochondrial stress have been carefully examined. As expected, the mammalian target of rapamycin (mTOR), a sensor of the mitochondrial transmembrane potential and ROS supply [55], is activated in lupus T cells [56]. The cellular growth program, in particular the manufacturing of proteins, depends on mTORC1 to sense nutrient availability, balance energy production and utilization, and direct energy carriers into proper pathways [57]. As part of this program, mTORC1 regulates the endocytic recycling machinery (Rab5A and HRES-1/Rab4 small GTPases), which determines the half-life and positioning of cell surface receptors, including the T cell surface receptor/CD3ζ chain (TCRζ) [56]. In lupus T cells, chronic mTORC1 stimulation sustains recycling and lysosomal degradation of TCRζ, impacting the sensitivity of such T cells to antigen stimulation. Compensatory upregulation of the Fcε receptor type Iγ chain (FcεRIγ) and recruitment of the tyrosineprotein kinase Syk then mediates enhanced calcium fluxing, prolonging T cells activation and amplifying clonal expansion [58] (Figure 2). mTORC1 hyperactivation has also been implicated in IL-4 and IL-17 production, negatively impacting the population of regulatory T cells [59]. The concept that mTORC1-imposed oxidative stress is a key pathogenic pathway in SLE has been translated into clinical medicine (Figure 2). Treatment with N-acetylcysteine, a precursor of glutathione, has been shown to successfully block mTORC signaling and improve disease outcomes in both mouse models and SLE patients [60,61]. Along the same line, treatment with rapamycin, an inhibitor of the mTOR pathway, has also been reported to have beneficial effects in the management of SLE patients and alleviated disease progression [62].
In contrast to the disease-promoting role of mitochondria in driving T cell hyperreactivity, recent data have linked SLE susceptibility to mitochondrial failure. Patients with SLE usually present with decreased levels of complement and individuals born with deficiency of the complement component 1q (C1q) are at high risk for SLE [63]. In a paradigm-shifting study, Botto and colleagues have mechanistically connected C1q deficiency and abnormal adaptive immunity in SLE. Specifically, they found that C1q, but not C3, is capable to restrain the response to self-antigens by modulating the mitochondrial metabolism in CD8+ T cells [64]. It has been known for two decades that C1q, when bound to its cognate cell surface receptor p32, translocates into the mitochondrial matrix, where it stimulates mitochondrial function [65]. While T effector cells rely mostly on energy production through glycolysis, T memory cells require mitochondrial energy supply to support their biosynthetic and bioenergetic needs. In the Botto et al study, lack of C1q disfavored the survival of long-lived CD8 memory T cells and instead promoted the differentiation of short-lived, pro-inflammatory CD8 T cells [66]. Consequently, C1q-deficient mice were prone to produce autoantibodies and develop renal inflammation. These data directly link C1q deficiency, insufficient mitochondrial stimulation and tissue inflammation. Notably, the effect of C1q appeared to be relevant to regulate CD8, but not CD4 function, a deviation from the traditional paradigm that assumes that the major drivers of autoantibodies are CD4 helper T cells.
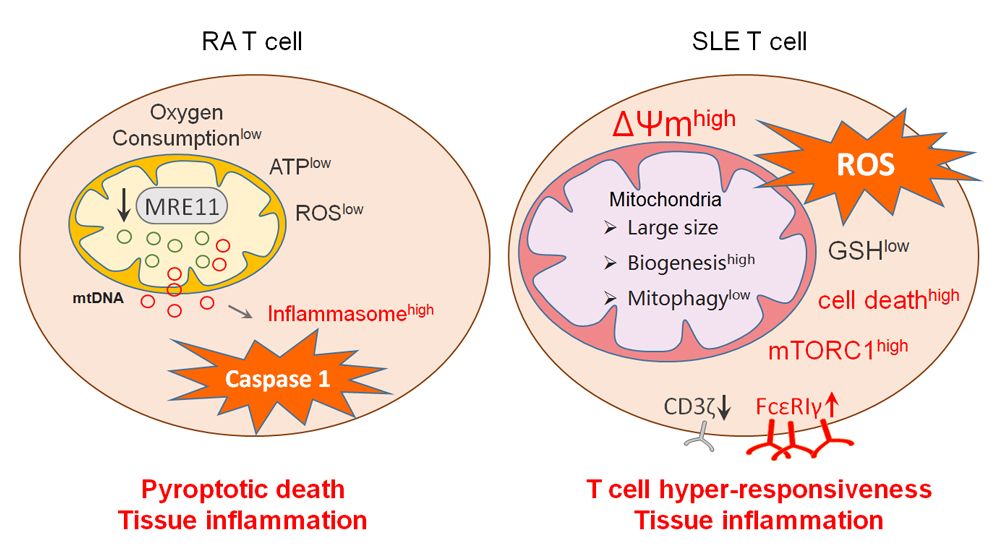 Figure 2. Mitochondria in RA and SLE T cells. RA T cells lose expression of MRE11, a nuclease functioning as a DNA repair molecule. Lack of mitochondrial MRE11A results in damage of mitochondrial DNA, increased susceptibility of mtDNA to oxidative damage and leakage of mtDNA into the cytoplasm. Cytoplasmic mtDNA is recognized as a danger associated molecular pattern (DAMP), initiates assembly of the inflammasome and activates caspase-1. As a result, T cells undergo pyroptotic cell death, release IL-1β and IL-18 and function as a nidus of tissue inflammation. In SLE T cells, mitochondrial biogenesis and fusion are enhanced while mitophagy is decreased. Thus, SLE T cells have high mitochondrial mass and elevated mitochondrial transmembrane potential [ΔΨm]. Inevitably, such T cells produce abundant ROS and exhaust cellular glutathione stores. The oxidative intracellular environment leads to cell death and drives mTORC1 activation. Downstream events of mTORC1 activation include recycling and lysosomal degradation of the T cell surface receptor/CD3ζ chain (TCRζ). Compensatory upregulation of the Fcε receptor type Iγ chain (FcεRIγ) and recruitment of tyrosine-protein kinase (Syk) enhance calcium fluxing, rendering these T cells hyper-reactive. MRE11, meiotic recombination 11; ROS, reactive oxygen species; GSH, glutathione; mTORC1, mammalian target of rapamycin complex 1.
Figure 2. Mitochondria in RA and SLE T cells. RA T cells lose expression of MRE11, a nuclease functioning as a DNA repair molecule. Lack of mitochondrial MRE11A results in damage of mitochondrial DNA, increased susceptibility of mtDNA to oxidative damage and leakage of mtDNA into the cytoplasm. Cytoplasmic mtDNA is recognized as a danger associated molecular pattern (DAMP), initiates assembly of the inflammasome and activates caspase-1. As a result, T cells undergo pyroptotic cell death, release IL-1β and IL-18 and function as a nidus of tissue inflammation. In SLE T cells, mitochondrial biogenesis and fusion are enhanced while mitophagy is decreased. Thus, SLE T cells have high mitochondrial mass and elevated mitochondrial transmembrane potential [ΔΨm]. Inevitably, such T cells produce abundant ROS and exhaust cellular glutathione stores. The oxidative intracellular environment leads to cell death and drives mTORC1 activation. Downstream events of mTORC1 activation include recycling and lysosomal degradation of the T cell surface receptor/CD3ζ chain (TCRζ). Compensatory upregulation of the Fcε receptor type Iγ chain (FcεRIγ) and recruitment of tyrosine-protein kinase (Syk) enhance calcium fluxing, rendering these T cells hyper-reactive. MRE11, meiotic recombination 11; ROS, reactive oxygen species; GSH, glutathione; mTORC1, mammalian target of rapamycin complex 1.
T cells are distinguished from other immune cell types by their enormous capacity to proliferate. The building of biomass is only possible through the well-tuned and dynamic utilization of energy sources and the proper assignment of energy carriers to either ATP generation or the production of biosynthetic precursors. Producing the membranes for intracellular organelles and the plasma membrane needed for daughter cells imposes high requirement for lipids. In the current paradigm, extracellular lipid supply is not considered a key element to drive uncommitted T cells towards lineage specification for Th1, Tfh or Th17 effector cells. However, intracellular lipid metabolism appears critical for pro-inflammatory T cell function, given the core role lipids play in the membrane systems. Support for the concept that fatty acids increase the propensity towards inflammatory activity comes from studies in obesity. Here, the metabolic stress induced by obesity has been reported to shift T cell differentiation towards the pro-inflammatory effector phenotype [67].
Compared to healthy T cells, RA T cells upregulate a broad, yet selective, lipogenic gene program focused on fatty acid (FA) synthesis. Acetyl-CoA carboxylase 1 (ACACA), fatty acid synthase (FASN), stearoyl-CoA desaturase (SCD), and the fatty acid–coenzyme A ligase family are critical enzymes in de novo fatty acid synthesis and elongation and all of them are upregulated in RA T cells as they differentiate into effector cells [68] (Figure 3). Fatty acid synthesis is further facilitated by the availability of NADPH produced in the PPP and high levels of intracellular acetyl-CoA [69]. Although mitochondrial fatty acid oxidation has not been investigated in detail, given the low mitochondrial activity and low ATP output [39], it is reasonable to predict insufficient fatty acid utilization in RA T cells. Declining lipolysis would obviously further increase fatty acid accumulation. In line with these data, both ex vivo-stimulated T cells and synovial T cells from RA patients deposit excess fatty acids as cytoplasmic lipid droplets (Figure 3). This metabolic deviation is closely linked to an upregulated locomotion program of RA T cells [68]. The movement of T cells migrating through extracellular space is distinct from that of myeloid cells. T cells utilize amoeba-like locomotion, constantly changing their shape as they edge along collagen fibrils. T cells do not degrade extracellular matrix and thus must squeeze through preexisting matrix gaps [70]. Thus, T cell motility depends on cellular polarization, in which the cytoskeleton is reoriented and dynamic cell protrusions are formed. Compared with healthy T cells, RA T cells are more efficient in entering the collagen matrix and are faster in penetrating into deeper layers. RA-specific changes in T cell locomotion have been successfully captured in transwell chambers [68]. Inhibition of lipogenesis has been reported to correct the hypermigratory properties of RA T cells and effectively suppresses synovitis in a human synovium-chimeric mouse model. It is not surprising that the T cell motility machinery is integrated with abnormal lipid metabolism, as the turnover of membrane structure in migrating T cells largely relies on lipid availability. Accordingly, in T cells with lipid droplet storage, plasma membranes built invasive membrane ruffles. Molecular studies have revealed that formation of membrane protrusions occurs through the co-localization of the cytoskeletal marker F-actin and the membrane marker cortactin [71,72], reminiscent of podosomes utilized by tumor cells to spread and metastasize [73] (Figure 3).
In this context, it is remarkable that rerouting of the glycolytic machinery will eventually alter lipogenesis. This is exemplified in RA T cells, which lack PFKFB3 and have a low yield of pyruvate, yet upregulate lipogenic enzymes [68]. Mechanistically, enhanced fatty acid synthesis was linked to induction of a gene module encompassing a set of motility-associated genes, including TKS5, a scaffolding protein localized at the plasma membrane and important for the formation of invasive membrane ruffles [74]. Upregulation of TKS5 transcription in RA T cells is a prerequisite for tissue invasiveness [68]. Thus, lipid metabolism is firmly integrated into disease-relevant effector functions.
Lupus T cells are more sensitive to T cell antigen receptor (TCR) stimulation, with a decreased threshold of activation required to induce intracytoplasmic calcium (Ca2+) fluxing [75]. Glycosphingolipids (GSLs) and cholesterol partner to form lipid rafts in plasma membranes and are important in regulating TCR signaling, as such specialized membrane structures support the formation of the T cell receptor signalosome [76,77]. Compared with healthy controls, CD4 T cells from SLE patients support increased synthesis of GSLs [78] (Figure 3). One possible mechanism is the elevation of the lipid-responsive nuclear receptor LXRβ, together with its target genes NPC1 and NPC2 (Figure 3). Increased GSLs internalization and recycling has been described for SLE CD4 T cells, supporting the building of plasma membranes with distinct lipid composition [78]. Inhibition of GSLs synthesis beneficially affected signaling defects and the hyperproliferative phenotype in T cells from SLE patients. Friend leukemia virus integration 1 (FLI1), an ETS family transcription factor, is upregulated in SLE T cells [79,80], and has also been reported to interfere with GSLs metabolism and TCR signaling (Figure 3). Cholesterol, on the other hand, has not been linked to autoimmune T cell functions. However, statins have been shown to regulate the Th17/Treg balance and could have beneficial effects in several autoimmune diseases including multiple sclerosis, RA and lupus [81].
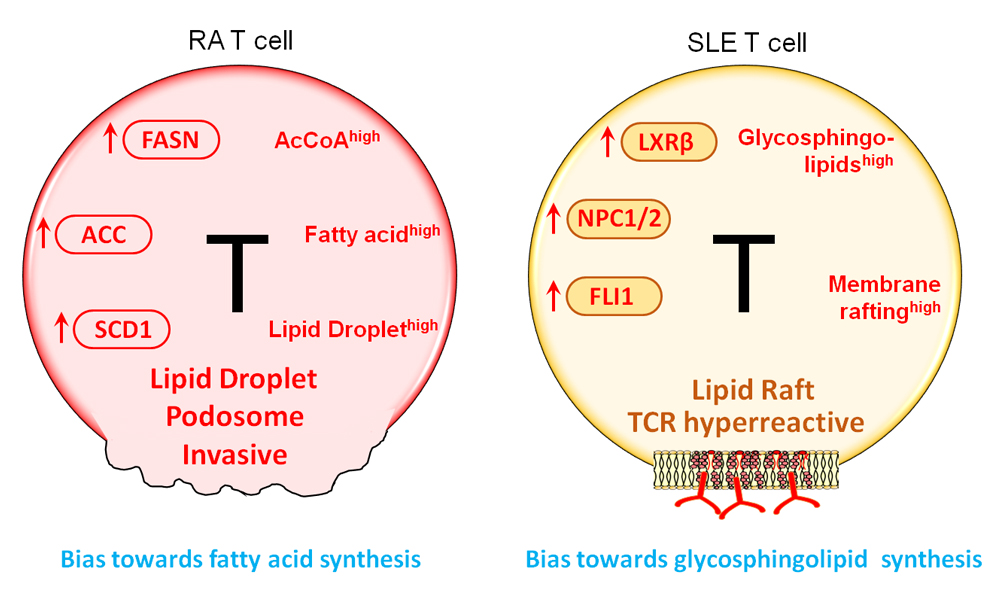 Figure 3. Lipid metabolism in RA and SLE T cells. Despite their ATPlow status, RA T cells activate the lipogenesis machinery. Enzymes supporting de novo fatty acid synthesis, including FASN, ACC, and SCD1 are all upregulated. Abundancy in intracellular AcCoA and NADPH facilitate the shift towards lipid generation. As a result, lipid droplets accumulate in the cytoplasm where they serve as a reservoir of biosynthetic activity. Equipped with surplus lipids, RA T cells tend to form membrane extensions; podosome-like structure that enable T cells to invade into matrix and promote tissue inflammation. In SLE T cells, glycosphingolipids (GSLs) synthesis is enhanced. Oversupply in GSLs changes the plasma membrane composition and structure, promoting lipid rafts formation. One outcome of membrane rafting is the enhancement of TCR signaling, enabled by easing the recruitment of lipid raft-associated signaling mediators, such as the proteins of the TCR complex and downstream signaling molecules. FASN, fatty acid synthase; ACC, acetyl-CoA carboxylase; SCD1, stearoyl-CoA desaturase; TCR, T cell receptor.
Figure 3. Lipid metabolism in RA and SLE T cells. Despite their ATPlow status, RA T cells activate the lipogenesis machinery. Enzymes supporting de novo fatty acid synthesis, including FASN, ACC, and SCD1 are all upregulated. Abundancy in intracellular AcCoA and NADPH facilitate the shift towards lipid generation. As a result, lipid droplets accumulate in the cytoplasm where they serve as a reservoir of biosynthetic activity. Equipped with surplus lipids, RA T cells tend to form membrane extensions; podosome-like structure that enable T cells to invade into matrix and promote tissue inflammation. In SLE T cells, glycosphingolipids (GSLs) synthesis is enhanced. Oversupply in GSLs changes the plasma membrane composition and structure, promoting lipid rafts formation. One outcome of membrane rafting is the enhancement of TCR signaling, enabled by easing the recruitment of lipid raft-associated signaling mediators, such as the proteins of the TCR complex and downstream signaling molecules. FASN, fatty acid synthase; ACC, acetyl-CoA carboxylase; SCD1, stearoyl-CoA desaturase; TCR, T cell receptor.
During their life cycle T cells need to adapt to changing tissue environments, periods of high proliferative stress, clonal expansion and clonal retraction, episodes of quiescence and differentiation into fast and highly functional effector cells. A key element in the adaptability of T cells is their highly effective sensing of intracellular energy resources and metabolic activities. The AMP-activated protein kinase (AMPK) serves as the main energy sensor in almost all eukaryotic cells [82]. Canonically, AMPK monitors AMP/ATP and ADP/ATP ratios and restores energy homeostasis by enhancing ATP-producing catabolic processes and inhibiting energy-utilizing anabolic activities. Besides directly measuring the availability of ATP, AMPK is capable of assessing the metabolic status through several nucleotide-independent pathways [82]. As a trimeric complex, AMPK consists of a catalytic subunit (α subunit) and two regulatory subunits (β and γ subunits). Whatever upstream signal or regulation is employed, AMPK activation is initiated by the phosphorylation of the α subunit, and AMPK activation has been tightly linked to the pool of membrane-associated AMPK, rather than unbound cytosolic AMPK [83–85].
An open question has been how T cells from RA patients, given their diminished glycolysis and low ATP production [86], monitor and adjust their proliferative and biosynthetic activities as well as their commitment to the production of effector molecules. Initial studies revealed that AMPK activity was significantly inhibited in RA T cells [87], despite the low ATP conditions and high cellular turnover. Essentially, the cellular rheostat connecting energy supply and utilization is no longer intact in these T cells. Inappropriately low mobilization of AMPK signaling did however provide an explanation for several features of RA T cells: reduced mitochondrial activity and reduced ATP synthesis; low intracellular ROS concentrations; shunting of glucose into the PPP instead of into pyruvate generation; inability to metabolize surplus lipids, and persistently high activation of mTORC1. In a series of molecular and cell fractionation experiments, the mechanism underlying defective AMPK activation has been solved [87]. Essentially, RA T cells maintain AMPK in the unbound, cytosolic pool and fail to translocate the protein to the lysosomal surface (Figure 4). The misplacement of AMPK is a consequence of insufficient N-myristoylation of the β-subunit of AMPK (AMPKβ). Lacking a myristate tail, AMPK cannot be recruited to the lysosomal membrane and RA T cells fail to form the v-ATPase-Ragulator-AXIN/LKB1-AMPK super-complex, where AMPKα-Thr172 is phosphorylated by LKB1 [88]. An important consequence of AMPK misplacement is the disruption of AMPK-mTORC1 cross-regulation. Lysosomal AMPK suppresses activation of co-localized mTORC1, shifting the cells towards anabolic and away from catabolic activity [88] (Figure 4). Thus, RA T cells fail to prevent mTORC1 activation and several functional activities have been assigned to persistence of mTORC1 activity [87] (Figure 4). In human synovium-mouse chimeras, blocking mTORC1 activity via rapamycin therapy was highly effective to suppress synovitis [87]. The molecular defect causing AMPK mis-trafficking towards the unbound cytosolic pool has been identified. Addition of the myristic acid lipid tail to AMPK requires the enzyme N-myristoyltransferase-1 (NMT1) [87]. In RA T cells, this transferase is suppressed through a post-transcriptional mechanism (Figure 4). Replenishing NMT1 in RA T cells rescued AMPK activation and corrected the pro-inflammatory properties of these T cells. Interestingly, besides modulating mTORC1 signal, recent studies have shown that activation of AMPK can limit JAK-STAT-dependent signaling pathways, a major driver of RA-dependent events [89].
Inappropriate activation of mTORC1 is a shared feature of RA and SLE T cells, compatible with the notion that distinct metabolic abnormalities can convene to promote disease-relevant immunity. In T cells from SLE patients, mTORC1 hyper-activation has been identified as the main driver of T cell dysfunction. In clinical studies, the mTOR inhibitor rapamycin has shown promise in ameliorating SLE disease activity; presumably, by correcting pro-inflammatory T-cell lineage specification [62,90]. While inappropriate mTORC1 activity in SLE T cells has been attributed to mitochondrial stress and excess production of mitochondrial ROS, the role of AMPK in controlling mTOR signaling has been insufficiently examined. The AMPK activation status has not been systemically studied in SLE T cells, but indirect evidence, such as the high level of catalytic metabolism, the enhanced mitochondrial biogenesis and the intensified mitochondrial respiration, would all suggest that AMPK is in hyperdrive. Metformin, a known activator of AMPK and a first-line pharmacological therapy for type II diabetes mellitus, has been tested as an immunomodulator in SLE [25,91], but more definitive data are needed to gauge the potential role of targeting AMPK in treating SLE patients.
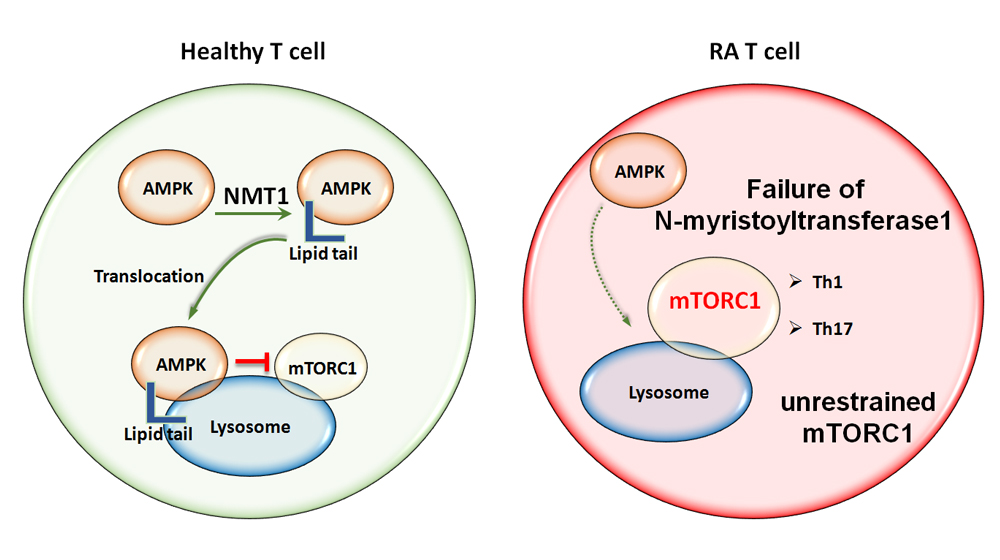 Figure 4. Misrouting of AMPK in RA T cells. AMPK is the major sensor of the cell’s energy status and co-ordinates energy reserves with biosynthetic activity by regulating mTORC1. AMPK activation occurs on the surface of the lysosome, where it co-localizes with mTORC1. Activated AMPK signals a need for catabolic activity and suppresses mTORC1. The recruitment of AMPK to the lysosomal surface requires the addition of a myristic acid lipid tail, that enables anchoring in the lysosomal membrane. Protein myristoylation is mediated by N-myristoyltransferase-1 (NMT1). Failure of NMT1 in RA T cells alters the intracellular trafficking of AMPK and prevents translocation to the cytoplasmic surface of the lysosome. One of the outcomes is unrestrained mTORC1 activation and a commitment of RA T cells to cellular proliferation and anabolic metabolism. AMPK, AMP-activated protein kinase; mTORC1, mechanistic target of rapamycin complex 1; NMT1, N-Myristoyltransferase-1.
Figure 4. Misrouting of AMPK in RA T cells. AMPK is the major sensor of the cell’s energy status and co-ordinates energy reserves with biosynthetic activity by regulating mTORC1. AMPK activation occurs on the surface of the lysosome, where it co-localizes with mTORC1. Activated AMPK signals a need for catabolic activity and suppresses mTORC1. The recruitment of AMPK to the lysosomal surface requires the addition of a myristic acid lipid tail, that enables anchoring in the lysosomal membrane. Protein myristoylation is mediated by N-myristoyltransferase-1 (NMT1). Failure of NMT1 in RA T cells alters the intracellular trafficking of AMPK and prevents translocation to the cytoplasmic surface of the lysosome. One of the outcomes is unrestrained mTORC1 activation and a commitment of RA T cells to cellular proliferation and anabolic metabolism. AMPK, AMP-activated protein kinase; mTORC1, mechanistic target of rapamycin complex 1; NMT1, N-Myristoyltransferase-1.
The recognition that the utilization of energy carriers and the biosynthetic activity for biomass generation are critical elements in determining cellular fate decisions and cellular differentiation has opened the emerging field of immunometabolism. Metabolic control of cellular behavior may be particularly important for inflammatory cells, such as autoreactive T cells that drive chronic tissue inflammation in autoimmune disease. T cells from patients with the autoimmune diseases RA and SLE display distinct metabolic signatures, and in both cases, metabolic deviations have been linked to abnormal cellular behavior (Figure 5). Accumulated data allow one to draw a metabolic landscape, in which metabolic interference may become a novel immunomodulatory therapy.
1.
2.
3.
4.
5.
6.
 Figure 5. Comparison of the metabolic signatures in T cells from RA and SLE patients.
Figure 5. Comparison of the metabolic signatures in T cells from RA and SLE patients.
The authors declare that they have no conflicts of interest.
This work was supported by the National Institutes of Health (R01 AR042527, R01 HL117913, R01 AI108906, R01 HL142068, and P01 HL129941 to CMW and R01 AI108891, R01 AG045779, U19 AI057266, and R01 AI129191 to JJG) and by the Encrantz Family Discovery Fund. VA MERIT Award I01 BX001669 from the United States (U.S.) Department of Veterans Affairs and with resources and the use of facilities at the Palo Alto Veterans Administration Healthcare System.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
Wu B, Goronzy JJ, Weyand CM. Metabolic fitness of T cells in Autoimmune Disease. Immunometabolism. 2020;2(2):e200017. https://doi.org/10.20900/immunometab20200017

Copyright © 2020 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




